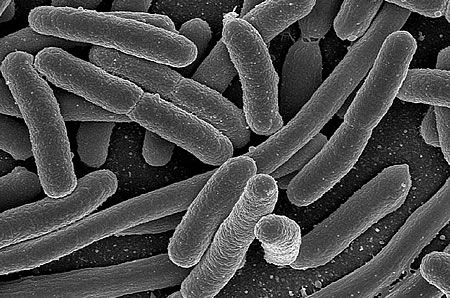
Bakterie (Bacteria, dříve též Bacteriophyta či Schizomycetes), nebo také eubakterie (Eubacteria), je doména jednobuněčných prokaryotických organismů. Mívají kokovitý či tyčinkovitý tvar a zpravidla dosahují velikosti v řádu několika mikrometrů. Studiem bakterií se zabývá bakteriologie, významně tuto vědu rozvinuli Robert Koch a Louis Pasteur.
Typickou součástí bakteriálních buněk je peptidoglykanová buněčná stěna, jaderná oblast (nukleoid), DNA bez intronů, plazmidy a prokaryotický typ ribozomů. U bakterií se nevyskytuje pohlavní rozmnožování, namísto toho se nejčastěji dělí binárně. Bakterie jsou nejrozšířenější skupinou organismů na světě. Dříve se druhy bakterií klasifikovaly podle vnějšího vzhledu, dnes jsou moderní zejména genetické metody. Díky nim se dnes rozlišuje asi 25 základních kmenů bakterií.
Bakterie mají velký význam v planetárním oběhu živin a mnohdy vstupují do oboustranně prospěšných svazků s jinými organismy. Mnohé patří mezi komenzálické druhy, které žijí například v lidské trávicí soustavě. Na druhou stranu je známo i mnoho patogenních bakterií, tedy druhů, které způsobují infekce. I člověk mnohé z bakterií využívá, například v potravinářském a chemickém průmyslu. Vědci využívají bakterie ve výzkumu.
Historie výzkumu bakterií
Bakterie poprvé pozoroval roku 1676 nizozemský přírodovědec Antoni van Leeuwenhoek, a to mikroskopem vlastní výroby. Jméno bacterium zavedl až Christian Gottfried Ehrenberg v roce 1838. Pojem pochází z řeckého slova bacterion, které znamená malý klacek či tyčka (první pozorované bakterie byly tyčinky). V roce 1859 Louis Pasteur dokázal, že kvašení způsobují bakterie, a že tyto bakterie nevznikají spontánně z neživé hmoty. Pasteur také prosazoval názor, že mikroorganismy včetně bakterií způsobují nemoci. Robert Koch byl průkopníkem v oblasti lékařské mikrobiologie a studoval původce cholery, TBC a anthrax. Při výzkumu TBC s konečnou platností dokázal, že bakterie jsou původci této nemoci, za což dostal v roce 1905 Nobelovu cenu. Takzvané Kochovy postuláty jsou výčtem čtyř kritérií, která jsou nutná k tomu, aby byl daný patogen uznán za původce určité nemoci.
V devatenáctém století již bylo známo, že jsou mnohé bakterie patogenní, ale nebyly známy účinné antibakteriální léky. V roce 1910 však Paul Ehrlich vyvinul první chemoterapeutikum proti bakterii Treponema pallidum (původce syfilis), a to díky záměně běžně používaných laboratorních barviv za sloučeninu, která selektivně zabíjela bakterie. Tradičně se však za první systémově použitelné antibiotikum uvádí penicilin, jehož účinků si všiml v roce 1928 Alexander Fleming. Gramovo barvení, metoda k rychlé klasifikaci bakterií do několika skupin, bylo vyvinuto v roce 1884 Hansem Christianem Gramem.
Spolu s vývojem bakteriologie se vyvíjelo i studium systematiky bakterií. Ve starším pojetí zahrnovala říše bakterie (Monera) všechny prokaryotní organismy. V roce 1977 Carl Woese zjistil, že Archaea mají vlastní vývojovou linii, odlišnou od linie bakterií. Z toho vyplývající taxonomie byla založená na sekvenaci určitého úseku rRNA, a rozdělila prokaryotické organismy na dvě samostatné domény.
Výskyt a životní nároky bakterií
Bakterie jsou nejrozšířenější skupinou organismů na světě. Celkově se odhaduje, že na Zemi žije asi 5×1030 (jedinců) bakterií. Celkový počet druhů se dá jen tušit, odhady sahají od 107 k 109 druhů. Bakterie je možné nalézt v půdě, vodě, ovzduší i jakožto symbionty uvnitř a na povrchu mnohobuněčných organismů. V jednom gramu půdy žije asi 40 miliónů bakterií, v jednom mililitru sladké vody je jich přibližně milion. Jsou však známy i druhy, které se specializují na prostředí, kde by ostatní organismy mohly přežívat jen stěží (vroucí voda v sopečných jezerech, nejvyšší vrstvy atmosféry a podobně). Některé druhy bakterií jsou dle výzkumů schopny přežít i ve vesmíru, tedy ve vakuu a o teplotě −270 °C.
Bakterie však mají různé požadavky na prostředí v němž žijí, významným hlediskem je teplota, kyselost a množství kyslíku. Podle teplotního optima se bakterie dělí na psychrofilní (do 20 °C), mezofilní (20-40 °C), termofilní (40 °C a více) a případně též hypertermofilní s optimem kolem 80 °C. Na základě kyselosti rozlišujeme alkalofilní (v zásaditém prostředí), neutrofilní (v ± neutrálním prostředí s pH 6-8) či acidofilní (v kyselém prostředí). Dalším významným hlediskem je vztah ke kyslíku. Aerobní bakterie (př. Mycobacterium) vyžadují kyslík v atmosférické koncentraci, mikroaerofilní (Lactobacillus) v koncentraci velmi nízké (cca 2 %), většina bakterií však patří mezi fakultativně anaerobní, které rostou lépe v přítomnosti kyslíku, ale dokáží růst i bez něho. Na okraji tohoto spektra jsou striktně anaerobní druhy, které žijí jen v prostředí bez kyslíku (většina druhů rodu Clostridium). Z dalších limitujících činitelů je možno zmínit vlhkost (většina je vlhkomilných, suchomilné jsou nokardie či aktinomycety), hydrostatický tlak (z hlubokých moří známe i barofilní bakterie), osmotický tlak (především v závislosti na množství solí), a podobně.
Tvar a velikost
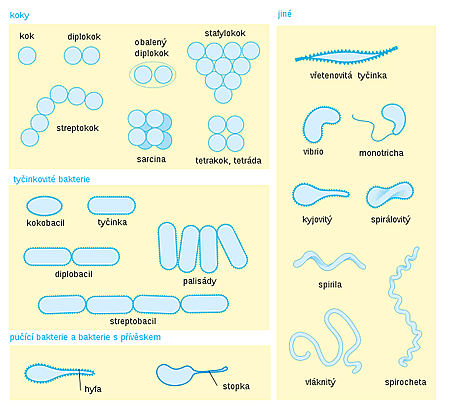
Navenek je nejnápadnějším rysem bakterií tvar bakteriálních buněk a jejich kolonií. U bakterií se rozlišují následující typy buněk dle tvaru:
- kulovitý (koky) — pokud vytvářejí kolonie, dělí se dále na diplokoky (kolonie tvořené dvěma buňkami), tetrakoky (čtyři buňky v kolonii), streptokoky (řetízkovité kolonie), stafylokoky (hroznovité kolonie) a sarciny (balíčkovité kolonie)
- tyčinkovitý (tyčinky čili bacily) — tyto se mohou sdružovat v koloniích po dvou (diplobacily) či v řetízcích (streptobacily), případně tvoří palisády
- zakřivený — takto tvarované bakterie nevytvářejí kolonie a patří mezi ně vibria (krátké lehce zakřivené tyčinky), spirily (lehce zvlněné tyčinky) či spirochéty (tyčinky šroubovitého tvaru)
- vláknitý — vláknité kolonie
- větvený — vytvářejí buďto náznaky větvení nebo větvení úplné. Druhá skupina může vytvářet bakteriální mycelia.
Některé bakterie vytvářejí kolonie podobné tělu jednoduchých mnohobuněčných eukaryot. Patří k nim některé sinice (např. Anabaena), myxobakterie (Myxococcales) a mnohé další skupiny. Druhé jmenované bakterie dokonce tvoří za určitých podmínek makroskopické plodničky se sporami uvnitř.
Rozměry bakterií
Velikost bakterií je velice rozmanitá, obvykle se ale pohybuje mezi desetinami a desítkami mikrometrů. Například Escherichia coli dosahuje délky 2–3 μm a šířky 0,6 μm. Je však známo mnoho v toto směru extrémních bakterií.
Dříve se mělo za to, že prokaryotická buňka nemůže být větší než řádově desítky mikrometrů vzhledem k omezeným možnostem difuze, která hraje hlavní úlohu při transportu látek buňkou. Mezi bakterie, které jsou zřejmě výjimkou z tohoto pravidla, patří například Epulopiscium fishelsoni, jejíž 0,2–0,7 mm dlouhé buňky jsou větší než buňky většiny prvoků. V současnosti je největší známá bakterie Thiomargarita namibiensis (0,75 mm).
Naopak nejmenší jsou bakterie rodu Mycoplasma (a příbuzný Ureaplasma) s průměrem buňky jen asi 0,1–0,3 μm. Nemají buněčnou stěnu, a proto byly dříve považovány za viry. Malé jsou i rickettsie a chlamydie. Ještě mnohem menší jsou však kontroverzní nálezy, označované jako nanobakterie, které dosahují velikosti jen 50 - 200 nanometrů (0,05 – 0,2 μm), podle nejnovějších výzkumů se však nejedná o živé organismy, ale o krystaly uhličitanu vápenatého v krevním séru.
Stavba buněk bakterií
Bakteriální buňka je buňkou prokaryotní. Přestože existují značné rozdíly mezi stavbou buněk různých skupin bakterií, lze vystopovat určité společné rysy. Známe nejen mnohé struktury na povrchu buněk (membrána, buněčná stěna, pilusy, bičíky), ale i vnitrobuněčné struktury (souhrnně protoplast).
Struktury na povrchu buněk bakterií
Na povrchu bakteriálních buněk je cytoplazmatická membrána podobná membráně eukaryot, ale s tím rozdílem, že většinou neobsahuje žádné steroidy. Důležitou funkcí membrány bakterií je tvorba ATP díky vytváření protonového gradientu (H+ iontů).
Většina bakterií má na povrchu buňky buněčnou stěnu z peptidoglykanu (murein), jenž obsahuje kyselinu muramovou jako jednu z hlavních složek. Peptidoglykan tvoří kolem buněk pevnou síť vyplněnou peptidy. Buněčná stěna bakterií je však velmi proměnlivá a je mimo jiné důležitým znakem při rozlišování bakterií na grampozitivní a gramnegativní. Grampozitivní mají totiž v buněčné stěně více peptidoglykanu, který zachycuje více krystalové violeti. Výjimku tvoří bakterie třídy Mollicutes (např. rod Mycoplasma), které nesyntetizují peptidoglykan a tudíž jim chybí buněčná stěna. Jejich tvar tak drží pouze třívrstevná membrána, do níž ukládají steroidy. Díky této zvláštnosti mají tyto baktérie velkou plasticitu a mohou měnit svůj tvar.
Mnoho bakterií má na povrchu ještě další polysacharidovou či proteinovou ochrannou vrstvu, tzv. kapsulu (pouzdro). Její funkcí je dále zpevňovat povrch bakterií (bakterie s kapsulou jsou díky tomu více patogenní) a navíc se lépe přichytit k substrátu. Někdy se díky slizovitým, méně soudržným kapsulám (zvaným slizová vrstva) vážou jednotlivé buňky v bakteriálních koloniích. Dohromady se kapsule a slizové vrstvě také říká glykokalyx.
Vyjma plošných útvarů, které kryjí buňku, se vyskytuje na povrchu ještě řada jiných struktur, jako jsou například bičíky a pilusy (rovněž zvané fimbrie). Bičíky jsou vlasovité (asi 20 nm silné, 20 µm dlouhé[8]) struktury ukotvené v membráně, tvořené helikálně složeným proteinem flagelinem a sloužící k pohybu. Bakterie mohou mít žádný, jeden (monotricha), více na jedné straně (lofotricha), dva uspořádané proti sobě (amfitricha) či velký počet bičíků rozložených po celém povrchu bakterie (peritricha). Bičíky bakterií se stavbou značně liší od bičíků eukaryot. Pilus čili fimbrie je druhým charakteristickým vlasovitým útvarem, ale kratší, tužší a užší v průměru. Pilusy bakterie využívají k přichycení na podklad (adheze), ale specializované sexuální pilusy (F pilusy) slouží k přenosu DNA (sexualita bakterií).
Protoplast
Uvnitř bakteriální buňky se nachází protoplast, tedy množství různých struktur a organel rozptýlených v cytosolu. Mezi nejvýznamnější vnitrobuněčné struktury u bakterií patří nukleoid (jaderné oblasti), ribozomy, inkluze a cytoskelet. V cytosolu však jsou rozptýleny i jiné struktury, například množství proteinů, mRNA a bakteriální metabolity.
Nukleoid (bakteriální chromozom, genofor) je jaderná oblast bakterií, tedy jediný obvykle kruhový řetězec tvořený dvěma vlákny deoxyribonukleové kyseliny (DNA). Vzácně je však DNA bakterií i lineární. Nejsou přítomny histony a netvoří se ani nukleozomy, celá oblast ani zpravidla není obklopena žádnou membránou. Opět však existuje výjimka a např. u planktomycet a poribakterií byly nalezeny membránové struktury, které připomínají eukaryotické buněčné jádro (u rodu Gemmata má dokonce toto jádro dvojitou membránu a póry). V cytoplazmě jsou pak roztroušeny plazmidy, malé úseky DNA. Ribozomy jsou prokaryotického typu; jsou charakterizovány zkratkou 30S+50S, která vyjadřuje velikost a strukturu ribozomálních podjednotek. Ribozomy bakterií jsou odlišné od eukaryotických nejen délkou proteinů, ale i jejich typem, čímž se logicky stávají častým cílem antibiotik. Inkluze jsou váčky či zrna, zpravidla sloužící jako zásobní struktury. Mohou skladovat glykogen, fosfáty, elementární síru, někdy však i barviva či enzymy.
Vznik a vývoj
Bakterie patří k nejstarším organismům nalézaných ve formě fosílií. První bakterie jsou známy z archaika, nalézány jsou stromatolity, tedy horniny obsahující fosílie bakterií, zvláště sinic.
Předkem dnešních bakterií byly jednobuněčné organismy, jež se vyvinuly na Zemi před čtyřmi miliardami let a patřily k vůbec prvním formám života. Další tři miliardy let všechny organismy na Zemi byly mikroskopické, přičemž po celou dobu dominovaly bakterie spolu s archea. Přestože byly objeveny fosílie bakterií (např. stromatolity), nedostatek zjistitelných znaků znemožňuje jejich určení a hlubší studium. Srozumitelné údaje se zjišťují pomocí sekvenace genomu recentních bakterií. Díky této technice vědci částečně rekonstruovali bakteriální strom života a zdá se, že bakterie jsou postranní větví linie, z níž se vyvinuly archea a eukaryota. Posledním společným předkem bakterií a archeí byl pravděpodobně hypertermofil, který žil před 2,5 – 3,2 miliardami let.
Klasifikace
Dnes se taxon bakterie (Bacteria) považuje za samostatnou doménu, která je striktně oddělena od ostatních (doména Archaea i všechny eukaryotické říše). Důvodem k tomuto rozdělení je skutečnost, že Archaea a Bacteria jsou velmi nepříbuzné skupiny lišící se velkým množstvím genetických i morfologických znaků.
Existuje více způsobů klasifikace bakterií. Zpočátku se klasifikovaly především podle vzhledu (fenotypu), dnes se však mimoto používají též analytické (podle chemických vlastností) a velmi často též genetické (podle genotypu) metody. Stejně jako u jiných organizmů se používá binomické názvosloví (jako je například Escherichia coli) a základním taxonem je druh. Nižšími taxony pak jsou poddruh, velmi často i morfovar, patovar a serovar.
Určování
Určování (determinace, identifikace) bakterií má velký význam v medicíně, kde je správným stanovením původce dané bakteriální infekce podmíněna následující léčba. Proto byla potřeba identifikovat tyto bakterie hlavním impulsem k vyvinutí determinačních technik. Mikroskopickým pozorováním tělních tekutin se bakterie určují jen zřídka, častěji jsou preparáty barveny. Známým typem barvení je Gramovo barvení, které umožňuje rozlišovat bakterie grampozitivní (G+), gramnegativní (G-) a bez buněčné stěny (Mollicutes). Pro mykobakterie (Mycobacteria) a nokardie (Nocardia) se zase používá Ziehl-Neelsenovo barvení. Často však nestačí ani barvit vzorek, ale přistupuje se ke kultivaci (viz níže). Při identifikaci bakterií také čím dál více používá také genetických metod, jako je polymerázová řetězová reakce. Jejich výhodou je jejich přesnost a rychlost v porovnání s kultivačními metodami.
Systematika
Systematika se zabývá pojmenováváním bakteriálních taxonů a jejich seskupováním podle příbuznosti. Klasifikace bakterií je průběžně vydávána v International Journal of Systematic and Evolutionary Microbiology (Mezinárodní ročenka systematické a evoluční mikrobiologie) a Bergey's Manual of Systematic Bacteriology (Bergeyho manuál systematické bakteriologie).
Dříve byla systematika založena především na základě morfologických a analytických metod, které však dnes vytlačují genetické metody (podobně jako v případě jejich určování). K používaným metodám patřilo i Gramovo barvení. Dalšími možnostmi bylo dělení na základě rozdílů v buněčném metabolismu, stavbě základních buněčných komponent (DNA, mastných kyselin, antigenů, a podobně). Tyto metody však nezaručují přirozenost taxonů, které byly na základě nich vytvořeny.
Dnešní bakteriální klasifikační metody se soustředí především na molekulární systematiku. Z genetických metod se využívá sekvenace dlouhodoběji stabilních částí DNA, jako je rRNA nebo je měřen obsah GC (množství guaninu a cytosinu v rámci DNA). Na základě zmíněných genetických metod je v rámci domény bakterie identifikováno poměrně velké množství kmenů. Jejich celkový počet není přesně dán a pohybuje se kolem 22 – 26. Příbuzné kmeny se seskupují do vývojových linií (klád), jedním ze známějších systémů je ten Cavalier-Smithův.
Růst a množení
Bakterie v prostředí, které jim zajišťuje pro ně vhodné chemické i fyzikální podmínky, zpravidla rostou a množí se. Mechanismem růstu je u bakterií syntéza všech komponent těla, čímž se zvyšuje hmotnost i objem jedince. Po dosažení určitých rozměrů se bakterie rozdělí metodou binárního dělení, což je typ nepohlavního rozmnožování. Doba mezi dvěma děleními se označuje jako generační doba. Při dostatečném počtu bakterií v dané populaci lze mluvit o kolonii, doba potřebná k zdvojnásobení počtu buněk v kolonii se nazývá doba zdvojení.
Nepohlavní rozmnožování
Bakterie se rozmnožují nejčastěji binárním dělením, je známo však několik případů odlišného typu nepohlavního množení (např. pučení, tvorba hormogonií, baeocyty a podobně).
Při binárním dělení se buňka nejprve prodlouží na dvojnásobnou délku a replikuje svou DNA, načež se uprostřed začne vytvářet septum (přehrádka složená z dvou membrán a základu buněčné stěny). Septum vždy vzniká růstem dvou přepážek od protilehlých stran buňky do jejího centra, kde se obě části spojí. Při tomto procesu se uplatňují různé enzymy (např. transpeptidázy). Běžně z každé mateřské buňky vznikne jedna sesterská buňka, při nedokončeném dělení septa však může dojít ke vzniku shluků bakterií (viz článek koky). U streptokoků jsou místa růstu vzájemně orientovaná pod úhlem 180°, čímž vznikají řetězce. U stafylokoků je tento úhel 90°, což má za následek vznik shluků.
Vyjma příčného dělení je známo ještě mnoho jiných alternativních způsobů nepohlavního rozmnožování. Zvláštním způsobem je například vytváření více než jedné spory, čímž se ze sporulace stává de facto rozmnožování. Takto se může za určitých podmínek dělit Anaerobacter (vzniká najednou až sedm spor). Další možností je mnohonásobné dělení jedné buňky na mnoho dílčích baeocytů u sinic řádu Pleurocapsales. Mateřská buňka obsahuje mnoho kopií DNA a v určitém okamžiku proběhne v rychlém sledu mnohonásobné dělení a z popraskané mateřské buňky se uvolní nové buňky. Naopak pučení, které se vyskytuje například u sinic nebo u Planctomycetes, probíhá tak, že zralá buňka začne na svém povrchu vytvářet zcela novou buňku. Pokud dceřiná buňka doroste do dostatečné velikosti, je do ní vpuštěna DNA a buňka (obvykle podstatně menší než mateřská) je odpojena. Postupem času doroste a může se začít sama rozmnožovat. I zde předchází pučení replikace DNA. Některé přisedlé bakterie (např. sinice) se množí pomocí hormogonií, krátkých vláken, která se oddělí od mateřského řetězce a dorůstají v nový.
Sporulace
Některé bakterie vytvářejí spory, tedy zvláštní buňky, které slouží k dlouhodobému přežití nepříznivých podmínek. Takové bakterie nazýváme sporulující. Spory tvoří zejména některé grampozitivní bakterie ze skupiny Firmicutes (modelové rody Clostridium či Bacillus). Jejich spory nazýváme endospory, protože vznikají uvnitř buňky mateřské, a to vždy jen jedna v každé buňce, zatímco okolní buňka mateřská se posléze rozpustí.
Jejich schopnost vydržet nehostinné podmínky se označuje jako kryptobióza. V praxi mohou spory přežít extrémně vysoké teploty (někdy vydrží až několik hodin varu), radiaci, kyselost prostředí, dezinfekční látky v prostředí a podobně. Tím se bakteriální spory pravděpodobně stávají nejodolnějšími známými buňkami v přírodě. Endospory se za příznivějších okolností opět změní ve vegetativní buňky.
Spory odlišného typu vytváří i jiné grampozitivní bakterie, konkrétně aktinomycety. Ty vznikají na konci vlákna aktinomycet, a proto se nejedná o endospory, a navíc mají mírně odlišné vlastnosti. Třetí skupinou, jež sporuluje, jsou bakterie rodu Azotobacter, v tomto případě se však odolné útvary spíše nazývají cysty. Cysty mají zpomalený metabolismus a ztlustlou buněčnou stěnu.
Kultivace
V laboratoři se mnohdy z různých důvodů bakterie kultivují v umělém prostředí (médiu). Toto prostředí musí mít vhodné chemické a fyzikální vlastnosti, zpravidla charakteristické pro určitý typ bakterií. Médium musí proto obsahovat všechny nezbytné živiny nutné pro život a růst bakterií. Každá bakterie požaduje určité minerální látky, ty se nejčastěji přidávají ve formě různých sloučenin.
Kultivační média prošla od dob Pasteura značným vývojem, jako živná půda pro mikroorganismy se používaly vývary z kvasnic, komorová voda z očí, později agar (polysacharid z ruduch). V klinické bakteriologii je dnes základem většiny živných médií krevní agar připravovaný smíšením ovčí krve a agaru. Může být dále upravován pro určité skupiny bakterií (např. čokoládový agar pro meningokoky, Šulova půda pro mykobakterie). Takzvaná selektivní média se používají v případě, kdy je potřeba získat čistou kulturu určitých bakterií - například po přidání 10% roztoku chloridu sodného vypěstujeme kulturu stafylokoků. Diagnostické médium umožňuje diagnostikovat určitý druh bakterií, např. podle jejich metabolického produktu.
Genetika
Většina bakterií obsahuje jediný nukleoid (jadernou oblast), obvykle kruhovou molekulu DNA, která se skládá z nukleových bází. Výjimkou jsou např. spirochéty z rodu Borrelia mající nukleoid lineárního (nikoliv kruhového) tvaru. Počet bází sahá od 160 000 komplementárních párů bází (u Carsonella ruddii) až k 12 200 000 párům u půdní bakterie Sorangium cellulosum. Geny jsou většinou po celé délce DNA, přestože i u bakterií existují introny (nepřepisované části DNA). Dále bakterie obsahují plazmidy, izolované části DNA, obsahující vždy jen několik málo genů. Bakterie se bez nich sice zpravidla obejdou, plazmidy však slouží bakteriím například pro rezistenci k antibiotikům i k jiným specializovaným účelům. Plazmidy se mohou přenášet mezi bakteriemi v rámci procesu konjugace.
Geny jsou v procesu transkripce přepisovány do jednotlivých typů RNA, jako je mRNA, rRNA či tRNA. Z mRNA se pak syntetizují proteiny v procesu translace. Bakterie se rozmnožují nepohlavně, a proto dědí identické kopie genomů svých rodičů (jsou to klony). Přesto se DNA může vyvíjet díky rekombinaci či mutacím Mutace způsobují chyby při replikaci DNA a vystavení různým mutagenům. Někdy může mezi bakteriemi docházet k výměně genetického materiálu.
Výměna DNA
Bakterie nemají systém pohlaví podobný eukaryotickým organismům, nicméně mohou si navzájem vyměňovat část svojí genetické informace. Tento proces se označuje jako konjugace. Přenos probíhá jedním směrem: jedna z bakterií je dárce čili donor DNA (nesprávně také „samčí buňka“), druhý je příjemce čili recipient („samičí buňka“). Výměna DNA se děje spojením buněk přes sexuální pilusy, vláknité duté útvary. Schopnost tvořit sexuální pilusy je umožněna specifickými sekvencemi DNA, jež se souhrnně označují jako F-faktor. V praxi může být F-faktor jak část bakteriálního chromozomu, tak plazmid. Dárce genetické informace musí mít F-faktor, proto je zván F+. Konjugací se zvyšuje genetická diverzita bakterií.
Bakterie mohou genetický materiál přijímat i z okolního prostředí, např. z mrtvých bakterií. Tento proces se označuje jako transformace. Proces transdukce zase v praxi znamená přenos genetického materiálu před bakteriální viry (bakteriofágy).
Metabolismus bakterií
U bakterií je známo velké množství různých metabolických procesů, pravděpodobně mnohem víc než u eukaryotických organismů. Tradičně byl typ metabolismu jedním ze základních znaků pro taxonomii bakterií, ale dnes se ví, že taxonomie založená na těchto znacích často neodpovídá skutečnému fylogenetickému vývoji. Metabolická aktivita je samozřejmě podmíněná obsahem biogenních prvků v substrátu (uhlík, dusík, síra, fosfor) a vhodného zdroje energie k biosyntetickým procesům. Auxotrofní druhy, tedy bakterie, které si neumí určitou sloučeninu syntetizovat, vyžadují též některé růstové faktory (vitamíny, aminokyseliny, purinové a pyrimidinové báze). Bakteriální metabolismus můžeme rozdělit podle tří kritérií: druh užívané energie, zdroj uhlíku a donor elektronů. Dle druhu užívané energie se rozlišují bakterie fototrofní a chemotrofní, podle zdroje uhlíku na autotrofní a heterotrofní a dle donorů elektronů na litotrofní a organotrofní. Tyto termíny jsou vzájemně kombinovatelné, proto například cyanobakterie jsou fotoautotrofní.
K fototrofním bakteriím patří především již zmíněné cyanobakterie (sinice), dále zelené sirné bakterie, zelené nesirné bakterie, heliobacterie, purpurové bakterie a v r. 2007 popsané chloracidobakterie. Většina se označuje za fotoautotrofy, protože zpravidla získávají uhlík z anorganických zdrojů (oxid uhličitý). U těchto bakterií probíhá fotosyntéza, proces, který pohání energie z slunečního světla a jeho výsledkem je fixace uhlíku v organických sloučeninách za pomoci Calvinova cyklu. Donorem elektronů pro redukci NADP+ je buď voda (tzv. oxygenní fotosyntéza, uvolní se kyslík), nebo jiné látky (vodík, síra, thiosulfát, sirovodík), při nichž se kyslík neuvolňuje. Sinice využívají při fotosyntéze barviva chlorofylu a dalších pigmentů. Mimo fotoautotrofy existuje i několik fotoheterotrofů, které sice také fotosyntetizují, ale jako zdroje uhlíku užívají organických sloučenin (acetát, pyruvát).
Chemotrofní bakterie, tedy bakterie využívající energie organických sloučeni, mohou být rovněž dvojího typu. Takzvané chemoautotrofní totiž získávají uhlík z oxidu uhličitého a chemoheterotrofní z organických sloučenin. K chemoheterotrofním bakteriím řadíme například rozkladače v půdě (v podstatě saprofyty) nebo bakterie podílející se na kvasných procesech. Známe však rovněž mnoho chemoautotrofních skupin, například bakterie vodíkové, sirné, bakterie oxidující železo či síru, nitrifikační a další.
Bakterie díky svým unikátním metabolickým pochodům hrají významnou roli v koloběhu látek, a to zejména v koloběhu uhlíku, dusíku a síry. Tím, že se podílejí na rozkladu organických látek, vrací do atmosféry oxid uhličitý. Některé bakterie dokáží z atmosféry fixovat vzdušný dusík, jako například Rhizobium, či sinice Anabaena a Nostoc. Některé z těchto bakterií jsou rovněž obsaženy v hlízkách bobovitých rostlin. Významné jsou i bakterie, zapojené v přeměně dusíkatých látek na dusitany a dusičnany, případně zpět na vzdušný dusík.
Ekologické vztahy
Bakterie jsou v přírodě nezastupitelné ve svém významu pro koloběh látek, jako symbiotické oboustranně prospěšné organismy či jako výrobní prostředek v biotechnologiích. Dále jsou tu také bakterie způsobující choroby a bakterie podílející se na rozkladu mrtvé organické hmoty (destruenti). Hydrolytické bakterie jsou zodpovědné za hydrolýzu organických látek.
Velmi významně se v historii bakterie zapojily do vzniku eukaryotických buněk. Bakterie mají i své patogeny, a to především viry, které se nazývají bakteriofágy.
Endosymbiotická teorie
Podle endosymbiotické teorie se dávné bakterie podílely na vzniku eukaryotické buňky z primitivních buněk, které mohly být příbuzné dnešním archeím. Alfaproteobakterie byly pohlceny protoeukaryotickými buňkami za vzniku mitochondrií či hydrogenozomů. Tyto organely se stále nachází u všech známých eukaryot (někdy v redukované podobě). Později vedlo pohlcení dalších endosymbiontů, tentokrát bakterií příbuzných sinicím, ke vzniku chloroplastů řas a rostlin. Existují dokonce řasy, jejichž chloroplasty vznikly pohlcením celé jiné řasy - jedná se o sekundární plastidy.
Mutualisté
Je známo velké množství mutualistických (oboustranně prospěšných) symbiotických vztahů bakterií. Velmi často se v symbiotických svazcích účastní sinice. Ty vstupují jak svazků s houbami (tzv. cyanolišejníky), ale velmi často též tvoří symbiotické vztahy s vyššími rostlinami (sinice jako fixátoři dusíku). Symbiotické sinice vegetující v tělech živočichů se nazývají zoocyanely a mají je například pláštěnci (Tunicata).
Mimo sinic je známo i mnoho heterotrofních bakterií, které se účastní symbiotických svazků. Významné jsou především hlízkové bakterie (např. Rhizobium) a také množství bakterií v tělních dutinách živočichů (například jako součást střevní mikroflóry). Známy jsou však i symbiotické bioluminiscenční bakterie, bakterie trávící celulózu a mnohé jiné.
Komenzálové
Na povrchu i uvnitř těl organismů žije poměrně značné množství bakterií, které nazýváme komenzálické, pokud svému hostiteli výrazně neškodí, ale ani výrazně neprospívají. V lidském těle se vyskytuje bohatá mikroflóra na kůži, v celé trávicí soustavě (zejména v tlustém střevě), v horních dýchacích cestách, v uchu a oku, močové trubici a vagíně.
Takzvaná střevní mikroflóra mnoha obratlovců včetně člověka je převážně tvořená právě komenzálickými bakteriemi, které jsou v tlustém střevě přítomny v obrovských počtech. Množství bakterií v gramu střevní tráveniny se odhaduje na 1012 a předpokládá se, že celkové množství střevních bakterií dokonce převyšuje množství lidských buněk. Někdy komenzálické bakterie přechází v patogeny, nebo se naopak stávají mutualisty.
Množství bakterií vytváří mikrobiální povlak i na lidské kůži. Přestože je lidská pokožka suchá, slaná a kyselá, roste na ní velké množství bakterií, jako Corynebacterium, Staphylococcus, Micrococcus a mnohé další. Většinou kožní bakterie žijí v blízkosti potních žláz a u kořene vlasů. Bakterie jsou i důvodem, proč lidé páchnou, když se potí.
Patogenní bakterie
Některé bakterie způsobují choroby, někdy souhrnně zvané bakteriózy či bakteriální infekce. Ty se neomezují na člověka, naopak různé bakterie napadají široké spektrum hostitelských druhů včetně hub, rostlin a prvoků. Přestože některé bakterie mohou být patogeny i na samotném povrchu těla daného organismu, značné množství jich vstupuje dovnitř různými tělními otvory, například u rostlin průduchy, skrz sliznice živočichů, ranami, případně přes kůži. V místě, kde se usídlí, mohou způsobit hnisání, ničit tkáň či pletiva (např. nekróza) či škodit svými vlastními toxiny.
Názvy bakteriálních nemocí se často tvoří přidáním koncovky -óza k názvu dané patogenní bakterie. Mezi nejběžnější lidské bakteriální nemoci patří například zubní kaz, z vážnějších nemocí je ve světě velmi častá tuberkulóza, v roce 2002 jí byly podle WHO infikovány dvě miliardy lidí a ročně na ní umíraly dva miliony lidí. Existují různé možnosti nákazy. S potravou (alimentárně) se do těla dostává například Salmonella (salmonelóza), Shigella spp. (shigelóza provázená průjmy) nebo Listeria spp. (původce listeriózy), vzdušnou cestou například Mycobacterium spp. (TBC a lepra) nebo Chlamydophila psittaci (původce jedné z chlamydióz). Ranami se do těla dostane například původce tetanu, Clostridium tetani. Speciální případ jsou přenosy přes členovce, vyskytující se například u bakterií Borrelia spp. (jeden druh původcem boreliózy), Rickettsia spp.(tyfus a jiné), a u mnohých dalších. Přes urogenitální trakt se bakterie mohou dostat do těla pohlavním stykem, jako například Neisseria gonorrhoeae (kapavka) či Treponema pallidum (syfilis).
Význam pro člověka
Využití v průmyslu
Bakterie, jako Lactobacillus, se (společně s kvasinkami a plísněmi) často již tisíce let používají k přípravě fermentovaných (kvašených) potravin, jako je sýr, sójová omáčka, nakládaná zelenina, kyselé zelí, ocet, víno a jogurt. Jogurt a kefír se vyrábí kvašením mléka za přítomnosti bakterií; mléko díky tomuto procesu dostává jiné příchuti. Mléčných bakterií se využívá při průmyslové produkci kyseliny mléčné. Ta totiž vzniká kvašením sacharidů. Fermentací sacharidů za přítomnosti máselných bakterií se zase využívá k produkci kyseliny octové a máselné, kvašením vzniklé kyseliny máselné se průmyslově vyrábí butanol a aceton. Dalšími látkami, jež jsou produkovány bakteriemi, jsou xanthan (používá se jako mazivo, přísada do potravin, při výrobě nátěrových hmot, keramiky a různých dalších prostředků) a kurdlany (potenciální uplatnění v potravinářství). V průmyslu se též uplatňují enzymy získané z bakterií. Některé proteázy se přidávají do některých pracích prášků, ke štěpení škrobu se užívá amyláz, v medicíně našly uplatnění streptokinázy.
Mnohdy se v biotechnologii přistupuje k cílenému šlechtění bakterií za účelem zlepšení jejich vlastností. V praxi to znamená hledat v kultuře náhodné mutanty, bakterie s, nebo bez určitého plazmidu, nebo je cíleně rekombinovat a mutovat. Uvažuje se například o šlechtění bakterií fixujících dusík.
Využití v ochraně životního prostředí
Schopnost bakterií rozkládat mnohé látky se využívá především v zpracovávání (např. toxického) odpadu i jiných způsobech bioremediace. V čistírnách odpadních vod je velmi často podporován růst aerobních rozkladných bakterií tím, že je odpadní voda promíchávána kyslíkem za vzniku tzv. aktivovaného kalu., rozkladných bakterií se však na podobném principu využívá i v různých domácích septicích.. Bakterie schopné trávit uhlovodíky v ropě jsou využívány při čištění ropných skvrn., na pláže se někdy přidává hnojivo, aby se růst bakterií podpořil (např. po havárii tankeru Exxon Valdez). V chemickém průmyslu se bakterie používají k produkci nejrůznějších chemických látek, případně léků či agrochemikálií. Bakterie se rovněž používají namísto pesticidů v biologickém boji proti škůdcům. V tomto ohledu je známá půdní bakterie Bacillus thuringiensis (BT).. Pomocí geneticky upravených bakterií se také vyrábí inzulin a další hormony, enzymy, růstové faktory či protilátky.
Využití ve výzkumu
Ve výzkumu se bakterií využívá kvůli rychlému růstu a poměrně snadné manipulaci s nimi. Bakterie jsou modelové organismy pro molekulární biologii, genetiku a biochemii. Vědci například cíleně mutují DNA bakterií a následné fenotypy zkoumají — tímto způsobem se zjišťuje funkce genů, enzymů a metabolických cest, jejich značná část se dá později aplikovat i na komplexnější organismy. Modelovým organismem je zejména bakterie Escherichia coli. Schopnosti některých patogenních bakterií inkorporovat plazmidy do DNA hostitele se využívá v genetickém inženýrství: zejména Agrobacterium tumefaciens je používaná při cílené přípravě geneticky modifikovaných plodin.
Kmeny bakterií
- Acidobacteria
- Actinobacteria - aktinobakterie
- Aquificae
- Bacteroidetes
- Chlamydiae - chlamydie
- Chlorobi
- Chloroflexi
- Chrysiogenetes
- Cyanobacteria - sinice
- Deferribacteres
- Deinococcus-Thermus
- Dictyoglomi
- Fibrobacteres
- Firmicutes
- Fusobacteria
- Gemmatimonadetes
- Nitrospirae
- Planctomycetes
- Proteobacteria - proteobakterie
- Spirochaetes - spirochety
- Thermodesulfobacteria
- Thermotogae
- Verrucomicrobia
K napsání tohoto textu byl mimo jiné použit materiál z wikipedia.org, diskuze.eu a sohu.cz.